


A növényszaporítás fajtái
Avagy a szaporítás ötven árnyalata

Eddig sokat beszéltünk már a növények szaporításáról, elsősorban a mikroszaporításról. Úgy gondolom, érdemes kicsit alaposabban belemenni a témába, így látni fogjuk majd, hogy a mikroszaporítás sem különbözik annyiban a hagyományos módszerektől, sőt: sok esetben csak egy kicsit felturbózott verziója. Továbbá kicsit bővebben szeretnék írni a folyamatokról, ami a mikroszaporítás szaporítás részénél történik.
Variációk
A különböző szaporodási formák ismerete azért fontos, mert a növénytermesztés során a növények természetes élettani folyamatait mesterségesen leképezzük és manipuláljuk. Tehát minél alaposabban tisztában vagyunk a háttérben zajló folyamatokkal, annál nagyobb sikereket tudunk elérni, illetve lehetőségünk nyílik új, innovatív megoldásokra.
A növényeknél két típusú szaporodást különböztetünk meg: az egyik a szexuális vagy generatív szaporodás, azaz magvetés, ekkor új genetikával rendelkező egyedek jönnek létre.
A magvetés akkor alkalmazható, ha az így létrejövő szaporulat megtartja a faj, fajta jellegzetességeit, vagy amikor hibrid egyedeket akarunk létrehozni. A hibridizáció két különböző csoporthoz1 tartozó egyed keresztezését jelenti (ilyen az előző hírlevélben már említett Anthurium × macrolobium, amely az Anthurium clarinervium és az Anthurium pedatoradiatum hibridje, ebből a keresztezésből találták az egyetlen darabot, amelyet Anthurium ‘Delta force’-nak neveztek el). A létrejövő utódok általában gyorsabban nőnek és nagyobb hozammal rendelkeznek, mint a szülők.
A mezőgazdaság - mind modern, mind korai formájában - elképzelhetetlen lenne hibridek nélkül: élelmiszer- és ipari növényeink vagy a termesztett fajtáik valamilyen módon hibridizáció eredményei. A hibridizáció természetes úton is előfordul, rengeteg kultúrnövényünk ősi formája így jött létre, például
az alma (Malus domestica)
a borsmenta (Mentha × piperita)
a földimogyoró (Arachis hypogaea)
a meggy (Prunus cerasus)
az arabika kávé (Coffea arabica)
és számos citrusféle (Citrus spp)
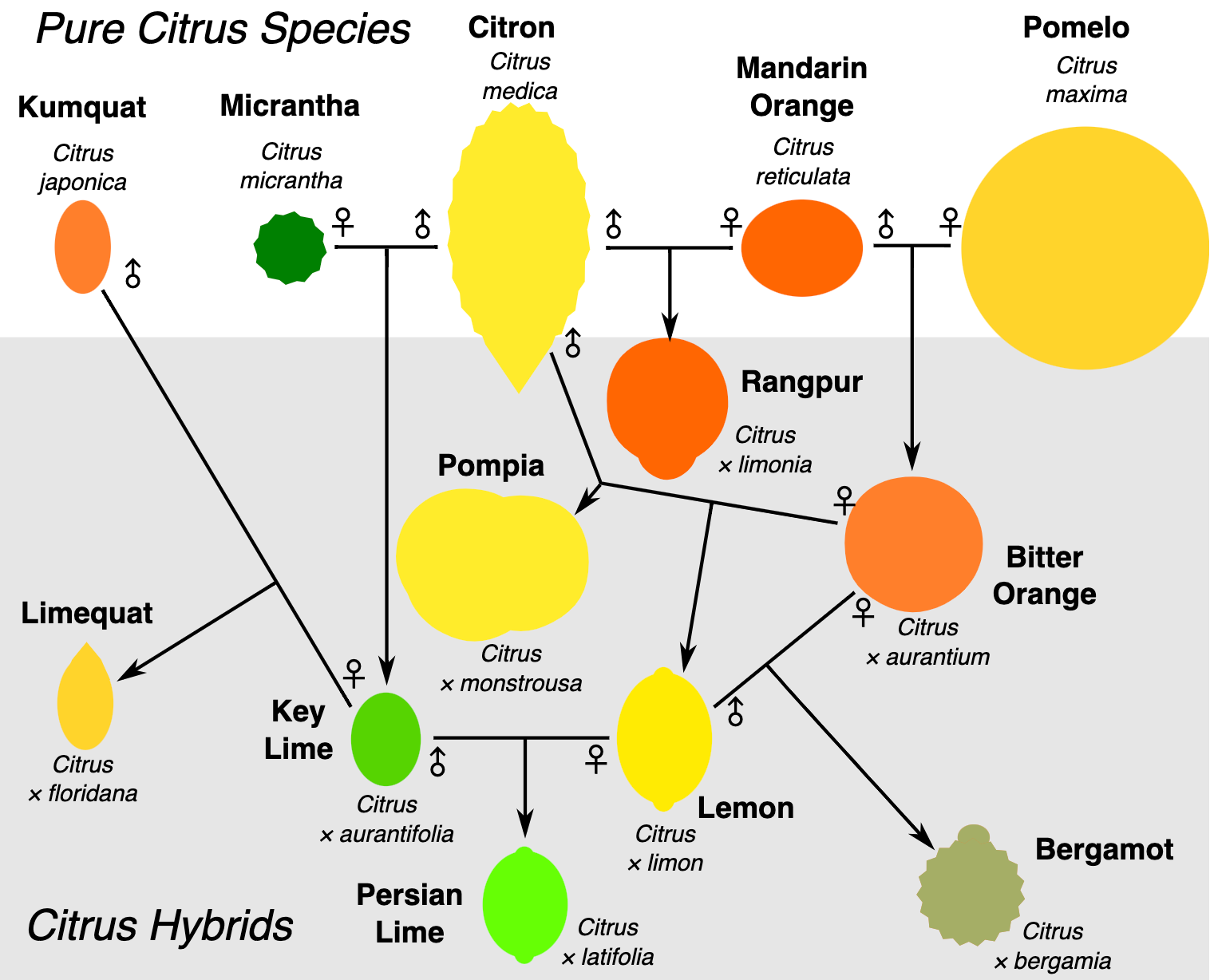
A (többnyire) mesterségesen előállított hibridekre pedig jó példa
a banán- és főzőbanánfajták (Musa × paradisiaca)
a durumbúza (Triticum durum)
a földieper (Fragaria × ananassa)
a paprika és paradicsom vetőmagok, stb.
Uniformitás
A másik típust aszexuális vagy vegetatív szaporodásnak nevezzük, és ebbe a kategóriába számos szaporodási forma tartozik. A vegetatív szaporodás folyamán az anyanövénnyel és egymással genetikailag azonos egyedek, úgynevezett klónok jönnek létre.
Ez növénytermesztés szempontjából kedvező, hiszen az adott fajra, fajtára jellemző tulajdonságokkal rendelkeznek, ugyanolyan ütemben növekednek, egységes alakot mutatnak, ám a genetikai változatosság hiánya komoly probléma forrása is lehet: mivel mindegyik növény ugyanolyan genetikával rendelkezik, egy esetleges vírus, bakteriális vagy gombás fertőzés akár az egész szaporulatot elpusztíthatja, mivel mindannyian ugyanannyira fogékonyak lehetnek a kórokozókra.
Erre a klasszikus példa a már említett banán: manapság 95%-ban a ‘Cavendish’ fajtát termesztik világszerte (ez talán a legsúlyosabb monokultúra a történelmünk során), ám az ‘50-es években egy másik fajta, a finomabbnak tartott ‘Gros Michel’ volt az egyeduralkodó. A váltást a Panama-betegségnek nevezett gombás megbetegedés okozta (Fusarium oxysporum f. sp. cubense), ami szinte kiírtotta az akkori banánültetvényeket az egész világon. A mai ellátási lánc rendkívül sérülékeny a monokultúra miatt, és komoly kutatások folynak egy esetleges katasztrófa megakadályozására.

A vegetatív szaporodás elsősorban az évelő, lágy- vagy fásszárú növényekre jellemző. Fiatalabb növényeket könnyebb vegetatívan szaporítani, mint idősebbeket, és az így szaporított növények gyorsabban elérik kifejlett formájukat a magról szaporítottakkal szemben. Ennek köszönhetően egy nagyon népszerű, olcsó és széles körben alkalmazott technológia már az idők kezdete óta.
A természetesen előforduló vegetatív szaporodási formák jellemzően módosult szárakból, levelekből vagy gyökerekből alakulnak ki. Ezek lehetnek:
inda (például eper)
sarjadzás (alma, banán, orchideák2, csokrosinda - Chlorophytum comosum)
ügynevezett elevenszülés (sarjika vagy elevenszülő korallvirág - Kalanchoe daigremontiana)
hagymák (tulipán, liliom, hagyma - Tulipa, Lilium és Allium)
gumók (burgonya, édesburgonya, dália - Solanum tuberosum, Impomoea batatas, Dahlia)
hagymagumók (kardvirág - Gladiolus)
apomixis vagy apogámia (lényegében aszexuális magképzés, számos növénycsaládban előfordul, például perjeféléknél (Poaceae), rózsaféléknél(Rosaceae))
Mesterséges vegetatív szaporítási formák:
dugványozás: az anyanövényről leválasztott szár vagy levél gyökereztetése, melyet kereskedelmi forgalomban kapható gyökereztető hormonnal lehet meggyorsítani
sarjleválasztás
bujtás és air-layering: a gyökereztetés az előtt történik, hogy a növényi részt az anyanövényről leválasztanánk
oltás: az alanyra (gyökér) egy nemest (rügy vagy szár) “ültetünk”, gyümölcsfáknál és zöldségnövényeknél különösen elterjedt
mikroszaporítás

A vegetatív szaporodási formák a növények nagyfokú regenerációs és gyökeresedési készségére hagyatkozik. Bizonyos esetekben nincs is különösebben feladata az embernek: a sansevieriák szívesen hoznak sarjakat a cserépben, ezeket csak le kell választani, vagy különösen a monsterák híresek hosszú léggyökereikről, ami akár különösebb előkészítés nélkül földbe ültethető a lemetszett növénydarabbal és hamarosan újabb példánnyal rendelkezünk.
A mikroszaporítás formái
A mikroszaporítás területén használunk magvetést és több vegetatív szaporodási formát is.
Magvetésre a legkézenfekvőbb példa az orchideák. Mint első hírlevelemben már említettem, a mikroszaporítás első igazi gazdasági mérföldköve az orchideák szaporításában elért siker volt.
Orchideákat szaporítani ugyanis lassú és körülményes dolog hagyományos eszközökkel, magvetésről pedig közel lehetetlen. Ennek az az oka, hogy az orchideák magja rendkívül apró, porszerű, nem rendelkezik úgynevezett endospermiummal, azaz tápanyaggal, amelyből a kis növény fejlődhet addig, amíg elég érett nem lesz a fotoszintézishez.
Hogyan lehetséges, hogy ennek ellenére a természetes környezetében léteznek, szaporodnak orchideák? A természetes élőhelyükön az orchideák speciális gombákkal állnak szimbiózisban: a szétszóródott apró magvak kapcsolatba kerülnek a gombafonalakkal, melyek ellátják tápanyagokkal őket, amíg kifejlődnek.
Mivel meghatározott gombafajokról van szó, melyek orchideafajonként eltérőek lehetnek, a magvetés szinte lehetetlen otthoni körülmények között a trópusi, egzotikus fajok esetén.3
A mikroszaporítás megoldást nyújt a problémára: mint már olvastuk, a táptalajban cukor és tápanyag található, amin ki tudnak fejlődni az apró növénykék. A steril környezet pedig lehetővé teszi, hogy ne kelljen megküzdeniük más mikroorganizmusokkal, amelyek biztosan elpusztítanák őket - azaz bepenészedne az orchideamag-vetésünk.

Így sem mondható gyorsnak a folyamat: 8-24 hónapba is beletelhet, amíg elég érettek lesznek ahhoz, hogy kivegyük őket az in vitro környezetből, és újabb évekbe telhet, amíg meghozzák első virágukat. A hagyományos orchidea nemesítés így egy különösen lassú és körülményes foglalkozás, a sokszor speciális környezeti igényekről nem is beszélve.
Mi történik az üvegben?
Az esetek többségében azonban aszexuális szaporodási formákat használnuk a mikroszaporítás során. A növényi rész, amit kultúrába vonunk, szinte bármi lehet: gyökér, szár, rügy, levél(nyél), virágzat.
A legegyszerűbb esetben egy rüggyel vagy hajtással kezdünk, amelyet hormonok segítségével manipulálunk. Amikor valamilyen szerv (például gyökér) az explantról közvetlenül fejlődik ki, azt organogenezisnek4 nevezzük. Gyökeresíthetünk in vitro, ami a hagyományos dugványozáshoz képest csak annyiban különbözik, hogy a steril, kontrollált környezet miatt jobb esélyekkel leszünk sikeresek, a növényt nem támadja meg például a penész, tehát nem rohad be, mint amikor egy pohár vízben áztatjuk.

A következő szint, amikor a rügyet vagy hajtást arra ösztönözzük, hogy az úgynevezett alvórügyeit is megnövessze, itt is újabb hajtásokat neveljen, amiket leválaszthatunk, és az előző bekezdésben leírt módon gyökereztethetünk, végül az új növényeket akklimatiizálhatjuk.
A komplexebb szinthez tartozik a merisztéma kultúra létrehozása. A merisztéma a még nem differenciálódott sejtekből álló szövet a növény különböző szerveiben. Ez azt jelenti, hogy a növények fejlődése során ez a szövet differenciálódik, válik valamilyen specifikus szövetté majd szervvé, például gyökérré, levéllé, kéreggé, virággá a megfelelő hormonok hatására. A merisztéma kultúra rügyekből vagy gyökércsúcsból szokás indítani.
A merisztéma-kultúra legnagyobb jelentősége a növények vírusmentesítésében van. A vírusfertőzés során az egész szervezet ferőzötté válik, így ezek a növények már nem használhatóak anyanövényként, nem lehet róluk szaporítóanyagot vételezni. Ez viszont komoly problémát jelent, ha csak a fertőzött növény rendelkezik a vágyott genetikával: fajának vagy fajtájának egyetlen ismert példánya, kívánatos, új mutációval rendelkezik, stb. A merisztéma-kultúra ebben az esetben megoldást nyújthat, ugyanis ez a szövete a növénynek az egyetlen, ahol vírusmentes marad. A merisztéma kinyerését már mikroszkóp segítségével végzik, annyira apró szövetről beszélünk (0,1-1mm nagyságrend).5
A mikroszaporításban gyakran használt módszer még az úgynevezett szomatikus embriogenezis6, ami annyit jelent, hogy egy új élőlény létrehozása ivarsejtek nélkül. Ebből tudjuk, hogy ismét egy vegetatív szaporodási módról beszélünk.
A szomatikus embriogenezis megértéséhez még egy szereplőről említést kell tennünk: a kalluszról. A kallusz a merisztémához hasonló differenciálatlan szövete a növénynek, melynek létrehozására a növényi hormonok megfelelő kombinációjával ösztönözhetjük a növényt.7

Ha kallusz nélkül jönnek létre az új növények, azt direkt embriogenezisnek nevezzük, míg amikor kalluszképződés után, differenciálatlan sejtekbből keletkeznek, akkor indirekt embriogenezisről beszélünk8. A növények ebben az esetben minden szervvel rendelkeznek, így megfelelő méret elérése után szétválasztjuk és akklimatizáljuk őket.
További fontos módszer még az embriókultúra: ennek során a fejletlen embriót helyezik in vitro környezetbe. Erre legtöbbször hibridizáció során van szükség, például a magnélküli fajták esetén, amikor az embriók (magok formájában) létrejönnek, de a termés érése során elhalnak és felszívódnak. A megfelelő időpontban kiemelt embrió azonban teljesértékű növénnyé nevelhető in vitro környezetbe helyezve (ebben az esetben embriómentésnek nevezzük).
Továbbiakban
Mint láthatjuk, egy olyan hétköznapi terület, mint a növények szaporodása, is tartogathat rengeteg érdekességet. A mai hírlevelemben igyekeztem összefoglalni és közérthetően elmagyarázni a különböző szaporodási / szaporítási formákat, valamint ennek segítségével még kézzelfoghatóbbá és érthetőbbé tenni, hogy mi történik a mikroszaporítás során.
A következő hírlevelemben a mikroszaporítás történetéről szeretnék írni: olyan sokszor került már szóba, hogy “már ekkor és akkor”, hogy úgy gondolom itt az ideje számba venni a nagyobb mérföldköveket.
Végezetül szeretném ismét megköszönni a nagy érdeklődést és a sok pozitív visszajelzést! Minden megosztás, üzenet és feliratkozás sokat jelent a számomra. Ha kérdésetek van, vagy lenne téma, amiről szívesen olvasnátok, hagyjatok kommentet!
A legelterjedtebb a fajok és fajták közötti hibridizáció, de akár ennél távolabbi fajok keresztezése is lehetséges. Dísznövények közt egy ismert példa lehet a rózsaszín vagy vörös szirmokkal rendelkező díszeper (Fragaria × Comarum), mely szülőfajai ugyanabba a családba (Rosaceae), de különböző nemzetségekbe (Fragaria és Comarum) tartoznak.
Az orchideák virágszárán vagy a növény tövénél megjelenő sarjakat keikinek nevezik.
Fontos hangsúlyozni, hogy nem minden orchidea esik ebbe a kategóriába: az orchideák családja több tízezer fajt számlál, az Antarktiszt leszámítva minden kontinensen megtalálhatóak, és ezért rendkívül változatosak. Ismertebb, magról nevelhető példa az Epidendrum radicans.
Az organogenezis a növények esetében folyamatosan történik, amíg életben vannak. Az új gyökerek és levelek is organogenezissel jönnek létre.
Ismét szeretném hangsúlyozni, hogy mennyire nem újdonságról van szó: Morel és Marthin már 1952-ben sikeresen létrehozott merisztéma-kultúrát.
Szomatikus: a testhez tartozó, tehát nem ivarsejt; embriogenezis: az embrió létrejötte.
Kallusz természetesen is létrejön a növényeken, például sebek felületén.
Steward és munkatársai 1958-ban, míg Reinert 1958-ban és 1959-ben sárgarépából nevelt kalluszon hoztak létre embriókat.